Chimie/Biologie: Des couleurs plein les yeux T2 Les acteurs de la coloration
|
|

La couleur dun corail est la résultante du mélange entre différentes couleurs, celles des pigments coralliens et celle à dominante brun doré des pigments contenus dans les zooxanthelles. Les densités respectives de ces deux composantes déterminent la couleur que nous percevons
(cf des couleurs plein les yeux T1 Lumière, couleurs et coraux).
En matière de coloration, tout est donc affaire de pigment, ce qui semble normal en regard de l'une des définitions du mot pigment : "substance naturelle colorée produite par les organismes vivants, en particulier végétaux", la pigmentation étant elle "la formation, l'accumulation de pigments dans les tissus vivants" ou encore "la coloration par un pigment" (© (c)Larousse).
La notion de pigment est donc suffisamment large pour englober des substances de nature et d'origine très différentes. Avant d'aller plus loin dans la compréhension des rôles respectifs des pigments présents dans les coraux photosynthétiques, il nous est donc apparu important de donner ici quelques points de repère sur ces familles de molécules.
Les pigments d'origine animale
Commençons ce rapide tour d'horizon en citant quelques molécules, qui bien que rentrant dans la définition donnée ci-dessus ne concernent pas directement notre hobby :
° L'hémoglobine est, bien sur, un pigment d'origine animale. Sa fonction principale est le transport de l'oxygène. Elle est composée d'une partie protéique, la globine, associée à un constituant non protéique, l'hème, qui contient un atome de fer ionisé (ion ferreux, Fe2+). C'est de l'hème que l'hémoglobine tire sa couleur rouge.
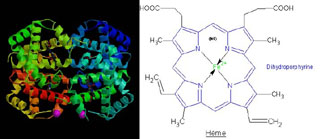
L'essentiel en chimie organique
° Les pigments biliaires sont des produits de dégradation de l'hémoglobine. Le plus important est la bilirubine qui joue notamment un rôle dans la digestion des graisses. Ils donnent à la bile stockée au niveau de la vésicule biliaire une couleur jaunâtre.
On est loin de la coloration des coraux nous direz-vous, et bien pas tant que ça, puisque l'hème dont il est question ci-dessus est très proche d'une structure que l'on retrouve dans la chlorophylle et dont nous reparlerons plus loin.
Il existe une autre classe de molécules produites par les animaux et qui, celle-là, a un rapport direct avec leur coloration puisqu'il s'agit des mélanines.
Deux types de mélanine existent. L'eumélanine, dont la couleur se situe entre le noir et le marron et la phémélanine qui est jaune-orangée. Ces pigments sont produits à partir d'un acide aminé - la tyrosine - dans des cellules spécialisées, les mélanocytes, situées dans les couches basales de l'épiderme. Si la mélanine est le principal responsable de la couleur de la peau chez l'homme, elle intervient également dans la coloration de l'épiderme de nombreux animaux et notamment des poissons.
Hémoglobines, pigments biliaires et mélanines sont les trois classes de pigments les plus fréquemment décrits lorsque l'on parle de pigments d'origine animale. Par origine animale, il faut comprendre "ceux qui sont synthétisés par les animaux". Ceci ne signifie pas qu'ils soient les seuls à être présents dans leurs tissus. Une classe très importante de pigments est en effet omniprésente chez les animaux sans que ceux-ci aient la possibilité de les synthétiser directement. Il s'agit des pigments caroténoïdes produits exclusivement par les végétaux, les algues et certaines bactéries. Ils ont donc chez les animaux une origine alimentaire, ceux-ci ayant la possibilité de les assimiler, voire de les transformer et de les utiliser pour des besoins spécifiques.
Les pigments d'origine végétale
Sont regroupés sous le terme de caroténoïdes de très nombreux pigments (plus de 300 répertoriés aujourd'hui). Il s'agit de terpènes, c'est-à-dire de molécules composées d'une chaîne d'atomes de carbone et d'hydrogène (comme le sont les hydrocarbures). Plus précisément, les caroténes et les autres pigments du même type sont composés d'une chaîne de 40 carbones dont les extrémités forment des cycles (voir figure ci-dessous).
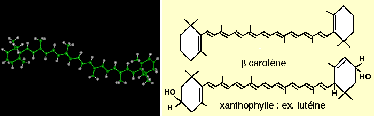
Les pigments caroténoïdes sont divisés en xanthophylles qui comportent des liaisons avec des atomes d'oxygène (O ou OH) et les carotènes qui n'en comportent pas.
Tous ces pigments sont qualifiés de pigments accessoires de la photosynthèse. Nous ne décrirons pas ici les détails des réactions mises en uvre lors de la photosynthèse. Mais, puisque c'est en grande partie grâce à celle-ci que nous arrivons à maintenir des coraux dans nos bacs (cf des couleurs plein les yeux Lumière, couleur et coraux), nous allons profiter de cet article pour rappeler quelques points marquants :
Tous les organismes photosynthétiques - les végétaux mais aussi certaines bactéries - sont capables de produire des composés organiques à partir de l'eau et du CO2 atmosphérique en utilisant l'énergie lumineuse. La réaction globale de la photosynthèse peut s'écrire :
n CO2 + nH2O + lumière -> (CH2O)n+ O2 (avec (CH2O)n = Glucides)
Les nombreuses réactions, mises en jeu lors de la photosynthèse, différent d'une espèce à l'autre mais comportent toujours deux catégories de réactions. Les premières permettent de convertir l'énergie lumineuse en énergie chimique en activant un électron de la chlorophylle. Celle-ci, devenue instable, cède alors cet électron à un 'accepteur' d'électron, puis retourne à son état d'origine en récupérant un électron sur une molécule d'eau ce qui a pour effet de séparer oxygène et protons :
2 H2O -> O2 + 4 H+ + 4 e-
La deuxième catégorie de réactions ne met pas directement en jeu la lumière. Dans cette étape, c'est l'énergie chimique produite précédemment qui est utilisée pour la transformation du CO2 en glucide :
CO2 + 4 H++4 e- -> CH2O + H2O
A l'issue de ces séries de réactions le bilan est donc bien :
2 H2O + CO2 -> CH2O + H2O + O2 et donc au final : H2O + CO2 -> CH2O + O2
Trois types de pigments peuvent être mis en jeu lors des processus de la photosynthèse dépendant de l'énergie lumineuse :
° Les chlorophylles (a, b et c) dont le rôle dans la photosynthèse vient d'être évoqué. Elles sont constituées d'une chaîne de 20 atomes de carbone rattachée à un noyau dont la structure est très proche de l'hème de l'hémoglobine (dans l'hémoglobine Mg est remplacé par Fe). Les chlorophylles a et b ne différent que par le remplacement, dans la structure II, du groupement CH3 par un groupement CHO (voir figure ci-dessous).
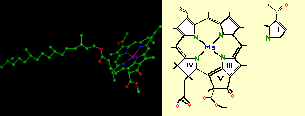
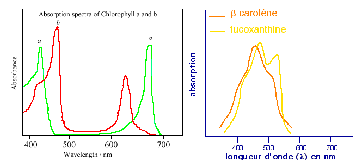
° Les pigments caroténoïdes sont qualifiés de pigments accessoires de la photosynthèse parce que l'énergie lumineuse qu'ils absorbent n'est pas directement utilisée mais est transférée aux molécules de chlorophylle avec lesquelles ils sont étroitement associés. Ils jouent néanmoins un rôle fondamental, puisqu'ils permettent d'élargir le spectre des longueurs d'onde utilisables pour la photosynthèse. Nous avons vu précédemment que la lumière était capable d'activer un électron de la chlorophylle. En réalité, seules certaines longueurs d'onde ont cette possibilité et correspondent, à peu de chose près, aux longueurs d'onde absorbées par la chlorophylle (les autres étant réfléchies). Les chlorophylles a et b ont des spectres d'absorption légèrement différents (cf figure ci-dessous) mais globalement elles absorbent les longueurs d'onde correspondant au bleu et au rouge et reflètent le jaune et surtout le vert (la couleur des feuilles, en lumière du jour, étant déterminée par ce spectre d'absorption). Avec seulement la chlorophylle, les longueurs d'onde situées entre 500 et 600 nm n'auraient donc pas d'action dans la photosynthèse.
C'est là qu'interviennent les différents pigments accessoires de la photosynthèse qui absorbent la lumière dans une plage de longueurs d'ondes plus large que les chlorophylles, et qui, même s'ils ne peuvent pas à eux seuls réaliser la photosynthèse sont capables d'activer les molécules de chlorophylle. La conséquence est donc pour les organismes qui en sont pourvu une meilleure utilisation du spectre solaire. A noter que ces pigments peuvent également jouer un rôle direct dans la chaîne de transport d'électron à la suite de l'activation de la chlorophylle. Enfin, ils semblent jouer chez certains végétaux un rôle photoprotecteur, notamment vis-à-vis des ultraviolets.
En raison de leurs différents spectres d'absorption, les caroténoïdes sont des pigments de couleur jaune, orange ou rouge. Ce sont eux qui donnent leur couleur aux feuilles d'automne lorsqu'à cette période la chlorophylle disparaît.
° La troisième catégorie de pigments intervenant dans la photosynthèse regroupe les phycobilines. Tout comme les caroténoïdes, il s'agit de pigments accessoires de la photosynthèse, mais qui sont présents essentiellement dans les algues et les bactéries.
Contrairement aux chlorophylles, les pigments caroténoïdes sont présents dans toute la chaîne alimentaire. Ils sont présents chez les animaux soit sous forme libre, soit associés à des lipides et/ou des protéines - caroténoprotéines ou caroténolipoprotéines. Ils sont à l'origine de la coloration de la plupart des invertébrés marins, notamment de la coloration des téguments des échinodermes, des mollusques et des crustacés, mais également des vertébrés qui les consomment(couleur rose de la chair du saumon). Chez les poissons, ces pigments alimentaires sont localisés, tout comme la mélanine, au niveau de cellules spécialisées de la peau - les chromatophores- et sont responsables de leur couleur.
Mais le rôle des caroténoïdes ne se limite pas à la coloration, ainsi par exemple, le béta carotène est le précurseur de la vitamine A.
Les pigments des coraux hermatypiques
Comme évoqué précédemment, les tissus des coraux hermatypiques hébergent des algues unicellulaires, les zooxanthelles. Celles-ci sont en fait des dinoflagellés du genre Symbiodinium présentes la plupart du temps au niveau du gastroderme des polypes (la deuxième couche de cellule des tissus coralliens, la première constituant l'épiderme). Les zooxanthelles étant photosynthétiques, les pigments caractéristiques des végétaux y sont présents, et notamment les chlorophylles a et c, des caroténoïdes et des phycobilines. La couleur brun-doré des zooxanthelles, est le reflet des différents spectres d'absorption de ces pigments (situés essentiellement au niveau des longueurs donde bleues/vertes).
Mais les coraux appartiennent au règne animal et les pigments caroténoïdes que l'on trouve dans leur tissus peuvent donc également être d'origine alimentaire. Chez les coraux non photosynthétiques, ce type de pigment ne provient que de l'alimentation. Les formes orange de Tubastrea par exemple, contiennent une très grande quantité de caroténoïdes qui sont vraisemblablement à l'origine de sa coloration si spécifique. Cette même couleur orange est due chez Montastrea cavernosa à des cynaobactéries symbiotiques contenant de très grande quantité de caroténoïdes. On ne sait pas encore si cette situation est rencontrée dans d'autres types de coraux (voir la news correspondante ).

Photo, http://www.photolib.noaa.gov
Certains pigments coralliens identifiés plus récemment appartiennent à une classe non encore évoquée ici, mais dont le rôle dans la coloration des coraux hermatypiques est prépondérant, les chromoprotéines. Ces pigments purement protéiques sont au cur de nombreux travaux de recherches depuis plusieurs années. Nous nous contenterons dans cet article d'en introduire les acteurs.
Pour la première fois en 1995, une équipe australienne (Dove et al. 1995) isolait le composant responsable de la coloration rose chez des pocilloporidés et des acroporidés. Dans les deux cas, il s'agissait d'une même protéine. Elle fut en premier lieu caractérisée chez Pocillopora damicornis et S. Dove lui donna le nom de pocilloporine. Des protéines similaires furent retrouvées aussi bien dans les variétés roses de Pocillopora, de Seriatopora hystrix ou de Stylophora pistillata que dans les formes bleues d'Acropora formosa ou digitifera. D'autres équipes ont depuis confirmé ces travaux, et retrouvé des protéines similaires dans toutes les espèces de coraux hermatypiques étudiées. La pocilloporine et les protéines de cette famille seraient donc responsables de la coloration rose, bleue, jaune mais aussi verte et brune (par mélange des précédents) de ce type de coraux.


Photo Etienne Dhont, Hervé Rousseau
Parallèlement à ces travaux, d'autres équipes travaillaient sur la fluorescence des coraux. Comme évoqué précédemment (Des couleurs plein les yeux T1 Lumière, couleur et coraux), la couleur que nous percevons dans le cas de la fluorescence, n'est pas due à la réflexion de la source lumineuse mais bien à la production de lumière par les pigments eux-mêmes.
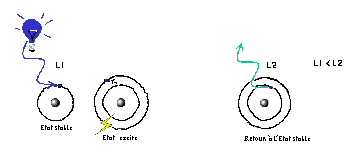
L'énergie fournie par la source lumineuse externe (ici sous la forme d'une radiation bleue) est captée par certains atomes des pigments. Ceux-ci passent alors d'un état stable à un état excité, correspondant au passage de leurs électrons d'une couche orbitale à une autre. Le retour des électrons à leur état initial fournit à son tour de l'énergie sous la forme d'une radiation de plus grande longueur d'onde que la source externe. D'où le passage, dans l'exemple ci-dessus, du bleu au vert.
Cette propriété de certains coraux et plus généralement des cnidaires (méduses, coraux et hydres) est décrite depuis de nombreuses années (Phillips, 1927 ; Catala, 1959). Curieusement, ce sont les travaux sur une méduse totalement transparente qui ont permis d'avancer dans ce domaine. Aequorea victoria est une petite méduse commune dans le Pacifique Nord à qui il arrive, à la suite d'une stimulation mécanique, d'émettre un flash de lumière verte. Deux protéines interviennent successivement au cours de ce processus, l'aequorine et la GFP (pour Green fluorescent protein, protéine fluorescente verte).
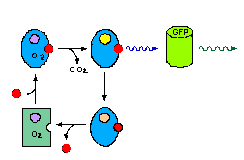
La fixation d'un ion calcium sur l'aequorine permet son activation et le passage à l'état excité de son chromophore (qui est le site qui, dans la molécule, est à l'origine de la coloration). Celui-ci revient à l'état stable en émettant une radiation bleue de 470 nm. Cette longueur d'onde est alors absorbée par la GFP qui en revenant à un état stable émet une radiation verte à 508 nm.
Chez Aequorea victoria, il y a donc à la fois bioluminescence - production de lumière en absence d'une source lumineuse externe - et fluorescence produite par la GFP.
Les travaux menés sur Aequorea victoria, ont incité certaines équipes scientifiques à rechercher et à découvrir des formes de la GFP chez d'autres coelentérés. C'est ce type de protéine (GFP-like) qui a pu être mise en évidence dans de très nombreuses variétés de coraux durs (acroporas), mous (discosomas, zooanthus) et même d'anémone (Heteractis crispa) et qui est responsable de leur fluorescence et de leurs étonnantes couleurs vertes et rouges sous lumière actinique.
Pocilloporines d'un coté, GFP-like de l'autre, deux familles de candidats sont donc en lice pour le titre de responsable de la panoplie de couleurs que nous cherchons tant à conserver dans nos bacs. Des études plus récentes ont permis de départager les candidats. Elles ont également apporté des éléments de réponse à la question qui servira de titre à notre prochain article :
Des couleurs plein les yeux Des pigments, mais pour quoi faire ?


Photos Hervé Rousseau
A suivre
Références
Catala, R. (1959) Fluorescence effects from corals irradiated with ultra-violet rays. Nature 183: 949.
S. Dove, M. Takabayashi, O. Hoegh-Guldberg (1995). Isolation and partial characterization of the pink and blue pigments of Pocilloporid and Acroporid corals. Biol. Bull. 189:288-297
Phillips, C.E.S. (1927) Fluorescence of sea anemones. Nature 119: 747.
Rechercher les contributions sur ce sujet dans le forum
Poser une question ou partager une expérience sur le forum
Article écrit par Benoit Finet et Florian Lesage et publié par Récifs.org le 16/01/05
|
| |
Score Moyen: 4.75
Votes: 12

|
|